 Glucógeno
Glucógeno
Los organismos
superiores se protegen de una posible escasez de energía mediante la
polimerización del excesos de glucosa y su almacenamiento como glucanos de alto
peso molecular (polisacáridos de a glucosa), que se pueden movilizar con
facilidad en momentos de necesidad metabólica. En las plantas a glucosa se
almacena como almidón, una mezcla del alfa-glucano, alfa-amilosa por la
presencia de ramificaciones. En los animales el glucano de almacenamiento es el
glucógeno, que solo se diferencia de la amilopectina en las ramificaciones.
 |
| Amilopectina |

El glucógeno se encuentra en gránulos citoplasmáticos que contienen hasta 120.000 unidades de glucosa. Su número es en especial importante en las células que usan grandes cantidades de glucógeno, el musculo (como máximo 1-2% de glucógeno en peso) y los hepatocitos (como máximo, 10% de glucógeno en peso, que corresponde a una provisión de energía ara el cuerpo.

Los gránulos de
glucógeno también contienen las enzimas que catalizan la síntesis y la
degradación del glucógeno, así como algunas otras enzimas reguladoras de estos
procesos.
¿Porque el
cuerpo realiza un esfuerzo metabólico tan grande para utilizar el glucógeno
como almacenamiento de energía si a grasa, que es mucho más abundante, puede
servir con el mismo propósito?
La respuesta tiene tres aspectos.
1.
Los músculos no pueden movilizar la grasa tan rápido
como lo hacen con el glucógeno
2.
Los residuos de los ácidos grasos de la grasa no pueden
metabolizarse en forma anaerobia
3.
os animales no pueden convertir los ácidos grasos en
glucosa, por lo que el metabolismo de as grasa por sí solo no puede mantener en
forma adecuada los niveles esenciales de glucosa en sangre.
Reserva de glucógeno
El organismo humano puede almacenar hasta 450 g de glucógeno;
una tercera parte de esta cantidad se localiza en el hígado y casi todo el
resto en la musculatura. El contenido de glucógeno de otros tejidos es escaso.
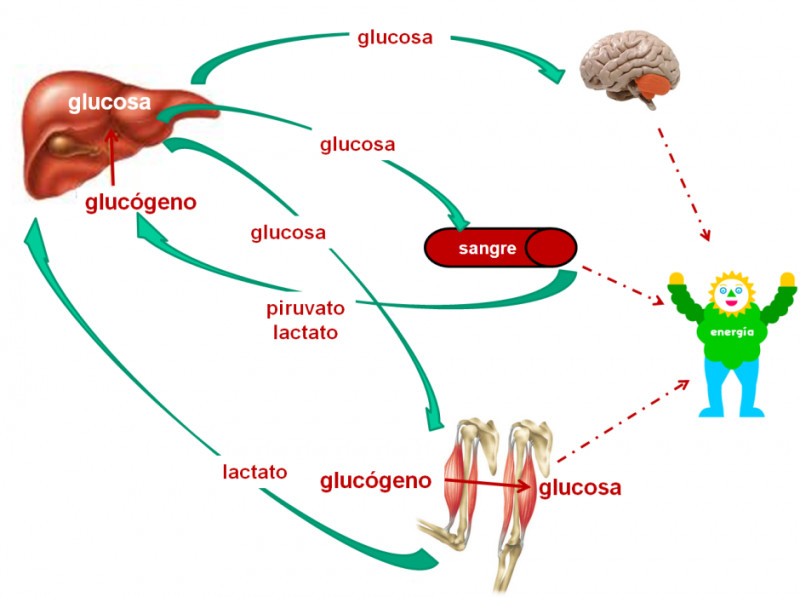
En la nutrición es de suma importancia el glucógeno, pues
esta sirve como base energética, cuando se realiza un ayuno prolongado, pues es
la primera reserva energética de glucosa. Esta reserva nos va a permitir estas
estables en cuanto a los requerimientos energéticos demandados por el
organismo.

Metabolismo del glucógeno
El glucógeno almacena la glucosa. La síntesis y la degradación del glucógeno se regulan con precaución para que pueda disponerse de suficiente glucosa para las necesidades energéticas del organismo.
La glucogénesis y la glucogenólisis están controladas principalmente por tres hormonas: insulina, glucagón y epinefrina.

´
Glucogénesis
La síntesis de glucógeno ocurre después de una comida,
cuando la concentración sanguínea de glucosa se eleva. Se sabe desde hace mucho
tiempo que justo después de ingerir una comida con carbohidratos ocurre la
glucogénesis hepática. La síntesis de glucógeno a partir de glucosa-6-fosfato
implica la siguiente serie de reacciones.
1. Síntesis de glucosa-l-fosfato:
La
glucosa-6-fosfato se convierte de forma reversible en glucosa-I-fosfato a
través de la fosfoglucomutasa, una enzima que contiene un grupo fosfato unido a
un residuo de serina reactivo:

El
grupo fosfato de la enzima se transfiere a la glucosa-6-fosfato, formando glucosa-l
,6-difosfato. Al formarse la glucosa-l-fosfato, el grupo fosfato unido a C-6 se
transfiere al residuo de serina de la enzima.
1. Síntesis de UDP-glucosa:
La formación de los
enlaces glucosídicos es un proceso endergónico. La formación de productos
derivados del azúcar con un buen grupo saliente proporciona la fuerza impulsora
para la mayoría de las reacciones de transferencia de azúcares. Por esta razón,
la síntesis de un nucleótido-azúcar es una reacción común que precede a la
transferencia de azúcar y a los procesos de polimerización. El difosfato de
uridina-glucosa (UDP-glucosa) es más reactiva que la glucosa y se mantiene de
forma más segura en el sitio activo de las enzimas que catalizan las reacciones
de transferencia (denominadas transferasas de glucosilo). Debido a que el
UDP-glucosa contiene dos enlaces fosfato, es una molécula muy reactiva. La
formación de UDP-glucosa, cuyo valor de t:,Go' es cercano a cero, es una
reacción reversible catalizada por la pirofosforilasa de UDP-glucosa:

Sin
embargo, la reacción se completa debido a que el pirofosfato (PP) es
hidrolizado de inmediato y de forma irreversible por la pirofosforilasa con una
pérdida grande de energía libre (G = - 33.5 kJ/mol):
3. Síntesis de glucógeno a partir de UDP-glucosa
La formación de glucógeno a partir de UDP-glucosa requiere dos enzimas: (a) de
la sintasa de glucógeno, que cataliza la transferencia del grupo glucosilo del
UDP-glucosa a los extremos no reductores del glucógeno, y (b) de la amilo-a- (1
,4--71 ,6)- glucosil transferasa (enzima ramificante), que crea los enlaces a (1,6)
para las ramificaciones de la molécula.
La
síntesis de glucógeno requiere de un tetrasacárido preexistente formado por
cuatro residuos glucosilo con enlaces a (1,4). El primero de estos residuos se
une a un residuo de tirosina específico en una proteína "cebadora"
que recibe el nombre de glucogenina. Después, la sintasa de glucógeno y una
enzima ramificante extienden la cadena de glucógeno. En el citoplasma de las células
hepáticas y en las musculares de animales bien alimentados pueden observarse
gránulos grandes de glucógeno, cada uno formado por una sola molécula de
glucógeno muy ramificada. Las enzimas causales de la síntesis y de la degradación
del glucógeno recubren cada gránulo

Balance global:
Glucosa-1-P + ATP + glucógeno + H2O ---> glucógenon+1 + ADP + 2
Pi
El glucógeno es una forma
muy eficiente de almacenamiento de glucosa, requiere poca energía: 1 ATP /
glucosa almacenada, si se parte de G6P o de G1P, si fuese desde glucosa libre
habría que invertir otro ATP.

Regulación de la
glucogenogénesis.
La actividad de la enzima
glucógeno sintasa es regulada por modificación covalente
(fosforilacióndefosforilación) en respuesta a la acción hormonal (adrenalina en
músculo y glucagón en hígado). La
glucógeno sintasa A (activa) es inactivada por fosforilación en un residuo
específico de SER a glucógeno sintasa B (inactiva) también llamada D, por ser
dependiente de la [G-6-P].
A la glucógeno sintasa A también se la llama I
(independiente)

Glucogenólisis
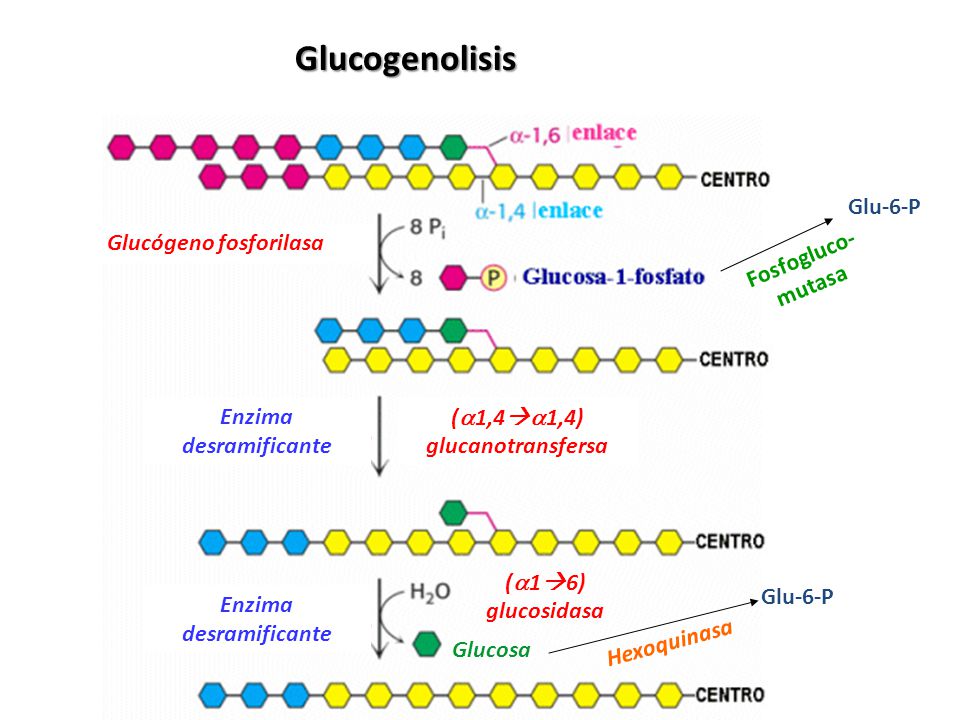
El hígado y el musculo son los tejidos principales de almacenamiento de glucógeno. En el musculo, la necesidad de ATP provoca la conversión del glucógeno en glucosa 6-fosfato (G6P) para ingresar en la glucolisis. En el hígado la baja concentración de glucosa en sangres desencadena la degradación del glucógeno en G6P que, en este caso, se hidroliza y se libera al torrente sanguíneo para revertir esta situación.
La degradación del glucógeno requiera la acción de tres enzimas:
1. Glucógeno fosforilasa (o simplemente
fosforilasa):
Cataliza la fosforolisis del glucógeno (ruptura del enlace por la
sustitución de un grupo fosfato) para producir glucosa-1-fosfato (G1P). Esta
enzima solo libera una unidad de glucosa que se encuentra como mínimo a cinco
unidades de distancia de un punto de ramificación.
2. Enzima desramificadora de glucógeno:
Elimina las ramificaciones del glucógeno, lo que permite completar la reacción de la glucógeno fosforilasa. La enzima también hidroliza las unidades glucosilo unidas mediante un enlace alfa (1—6) para producir glucosa. Como consecuencia, 92% de los residuos de glucosa del glucógeno no se convierten a G1P. cerca del 8% restante, que se encuentra en los puntos de ramificación, se convierte a glucosa.
3. Fosfoglucomutasa:
Convierte G1P a G6P que, también se forma en el primer paso de la glucolisis por la acción de la hexociasa o de la glucocinasa. El G6P puede continuar a lo largo de la vía glucolítica (como ocurre en el musculo) o hidrolizarse a glucosa (como en el hígado).
Regulación
de la glucogenólisis:
La glucógeno fosforilasa
es la enzima reguladora y está regulada mediante dos mecanismos:

a)
Regulación alostérica por metabolitos: EN
MUSCULO el AMP y EN HIGADO la glucosa.
b)
Modificación covalente reversible, por
fosforilación-defosforilación, como respuesta a la acción hormonal
Aquí ya hay que
considerar que la regulación del metabolismo glucídico es muy diferente en
músculo y en hígado. En el músculo el objetivo de esta vía es la degradación de
glucosa para la producción de ATP para la contracción y en el hígado cumple
otras funciones, mantener un nivel de glucosa constante en sangre; para lo cual
la moviliza desde el glucógeno y la exporta, o bien la importa y la almacena en
forma de glucógeno, para cuando la necesita; e incluso la produce.
A.
Modificación covalente de las
enzimas: mediante una CASCADA DE FOSFORILACIONES en respuesta a la acción hormonal:
(adrenalina en músculo y glucagón en hígado)
Existen dos formas de la
enzima que degrada el glucógeno, glucógeno fosforilasa a (R, fosforilada y
catalíticamente muy activa) y fosforilasa b (T, defosforilada y normalmente
inactiva). La fosforilación en un resto de SER de cada subunidad de la fosforilasa
b hace que se convierta en la fosforilasa a, y esa fosforilación la cataliza la
fosforilasa b quinas. La fosforilasa b quinasa se activa a su vez, por fosforilación
(también por alto nivel de Ca2+ en músculo). La enzima que cataliza esta última
fosforilación, de la fosforilasa b quinasa, es la proteína quinasa, que a su
vez se activa por la unión del AMPc
El AMPc se forma por la
adenilato ciclasa, en respuesta a la acción hormonal de la adrenalina en
músculo y del glucagón en hígado. Luego la cascada de fosforilaciones y activaciones
es:
HORMONA:
Adrenalina
Adenilato
ciclasa AMPc
Proteína
kinasa A
Fosforilasa
kinasa glucógeno fosforilasa a

Función del Glucógeno
Músculo: la glucosa se
degrada en la glicólisis para obtener ATP.
Hígado: se libera glucosa
a la sangre para mantener su nivel.

Regulación conjunta de
las dos vías: Glucogenogénesis y Glucogenólisis
El metabolismo del
glucógeno es regulado cuidadosamente para evitar el derroche de energía. Tanto
la síntesis como la degradación son controladas por un mecanismo complejo en el
que participan la insulina, el glucagón, la epinefrina y reguladores alostéricos.
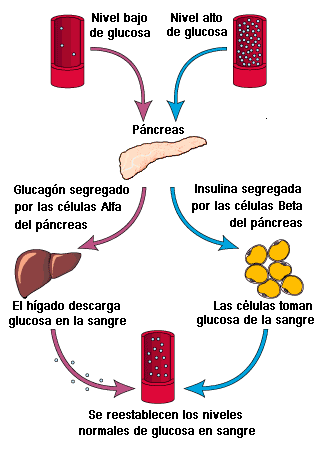
El páncreas libera glucagón cuando la glucemia decae en los periodos postprandiales.
Se une a receptores en los hepatocitos e inicia un proceso de transducción de
señales que eleva las concentraciones intracelulares de cAMPo El segundo mensajero,
el cAMP, amplifica la señal original del glucagón e inicia una cascada de fosforilación
que conduce a la activación de la fosforilasa de glucógeno junto con varias otras
proteínas. En segundos, la glucogenólisis provoca la liberación de glucosa en
el torrente sanguíneo.
Cuando está ocupado, el
receptor de insulina se convierte en una enzima cinasa de tirosina activa que
produce una cascada de fosforilación, la cual en última instancia tiene un
efecto opuesto al del sistema glucagón/cAMP: las enzimas de la glucogenólisis se
inhiben y las enzimas de la glucogénesis se activan. La insulina aumenta también
la velocidad de la introducción de la glucosa a numerosas clases de células diana,
pero no al interior de las células hepáticas o de las cerebrales.

La actividad de la sintasa
de glucógeno es estimulada por la glucosa-6-fosfato. En los hepatocitos, la
glucosa es un regulador alostérico que promueve la inhibición de la fosforilasa
de glucógeno.
Bibliografías
Mckee T., Mckee J., Metabolismo del glucógeno; Las bases
moleculares de la vida; México: D.F.; McGrawHill; 2014; Pág. 296-303
Koolman J., Rohm K., Metabolismo del glucógeno; Bioquímica;
Alemania; [Traducción] Editorial Panamericana; 2004; Pág.: 156.158
Voet D., Voet J., Metabolismo del glucógeno; Bioquímica;
Uruguay; Editorial Médica Panamericana; 2006 Pág.: 647-654